
论文:全州禾花鲤体质量和体长遗传参数估算及其选育进展
全州禾花鲤体质量和体长遗传参数估算及其选育进展
杜雪松,陈 忠,文露婷,潘贤辉,黄 姻,周康奇,林 勇,李 哲,罗 辉,覃俊奇
1. 广西壮族自治区水产科学研究院/广西水产遗传育种与健康养殖重点实验室,广西 南宁 530021
2. 西南大学 水产学院,重庆 402460
鲤 (Cyprinus carpio) 是人类最早驯化的鱼类,其经济性状的遗传改良对于满足人类蛋白质需求具有重要意义。中国拥有丰富的鲤种群和选育品系,如黄河鲤、荷包红鲤、瓯江彩鲤、松浦镜鲤等[1]。此外,分布在不同地域的稻田养殖系统也驯化和保存了多样性丰富的鲤鱼养殖品系[2],如青田田鲤、全州禾花鲤、金边鲤、瓯江彩鲤、乳源石鲤等[3-6],关键生产性状的遗传多样性丰富、改良潜力大,对其经济性状开展改良选育有利于促进养殖产业发展。目前,关于鲤生长性状的改良已开展了广泛的研究和实践,包括遗传评价体系、分子标记辅助选择 (Molecular-assisted selection, MAS)、全基因组选择 (Genomic selection, GS) 等[7]。其中,采用在遗传增益获得和群体近交水平控制方面具有明显优势的家系选育法,结合基于系谱信息的BLUP育种值估计选择,已选育获得了建鲤、福瑞鲤等新品种[8],为养殖产业做出了重要贡献。近年来,对分子标记辅助育种和全基因组选择育种技术的研究表明,其在生长性状改良选育种中具有更高效的改进效果,选择准确度比家系选育法高18%[7],是改良养殖品种经济性状的有效途径;然而,除乳源石鲤已获得“乳源1号”选育新品种之外[9-10],目前对稻田养殖鲤群体的改良选育报道极少,明显落后于黄河鲤、松浦镜鲤等。
全州禾花鲤 (C. carpio var. Quanzhounensis) 是广西北部稻鱼共生系统中驯化和保存的特色鲤鱼养殖品系,在当地称为禾花乌鲤或禾花鱼[11],品质极佳,曾作为贡品享有极高知名度。2010年,全州禾花鲤通过了原农业部农产品地理标志登记保护,是当地发展稻渔综合种养,提高稻田单位产出和综合效益的主要养殖品种,对乡村产业发展具有重要意义[12]。近年来,全州禾花鲤已被广东、湖南、贵州等省引进养殖,具有发展为全国性养殖品种的潜力。然而,全州禾花鲤生长速度较慢,早期测定的1龄鱼平均体质量为4.0~13.5 g、2龄鱼平均体质量为665~1 250 g[11],生长速度远低于建鲤、黄河鲤等,因此对该群体的生长性状改良已成为比较紧迫的产业需求。2017年,笔者团队针对全州禾花鲤生长速度慢的不足,启动了对其生长速度的改良选育工作。本文利用2017—2020年全州禾花鲤的选育改良数据,分析了其体质量和体长性状的表型参数,估算了相关遗传参数和选育进展,为开展稻田养殖鲤鱼群体生长性状的改良提供参考资料。
1 材料与方法
1.1 育种场地
本项目在广西农业良种南繁基地开展,该基地位于海南省乐东黎族自治县九所镇,年平均气温25.7 ℃,气温最高月为6月,平均28.7 ℃;气温最低月为1月,平均21.4 ℃。全年日照时间2 534 h,年平均降水量1 347.5 mm。
1.2 全州禾花鲤种质的引进
全州禾花鲤种群由桂林绿淼生态农业有限公司从桂林市全州县才湾镇13户农户的禾花鲤养殖稻田中收集;经过初步筛选后空运至广西农业良种南繁基地,共收集雄性321尾,平均体质量为95.58 g,平均体长133.0 mm;雌性251尾,平均体质量为32.84 g,平均体长88.70 mm;培育4个月后用于家系构建。
1.3 选育家系的构建
1.3.1 G0代家系构建
从收集到的全州禾花鲤种群中,随机挑选1尾雌性、1尾雄性进行1对1配对繁殖,产生全同胞家系。由于引进的禾花鲤亲本为农户稻田中的个体,养殖条件差异大、缺乏生产和生长时间记录,难以评估收集亲本的生长性状,因此G0代的亲本配对采用随机选择的方式进行。
1.3.2 G1—G2代家系构建
G1—G2代家系分别由上一代家系中选择的优秀个体经过1对1交配产生。各世代家系数量控制在30个以上,对照家系控制在5个以上 (各世代建立的家系数量见表1);优秀个体的选择采用体质量的育种值进行排序,在各家系中选择育种值排名最前的3~5尾作为候选亲本;对照家系的亲本选择育种值接近总体平均值的个体。
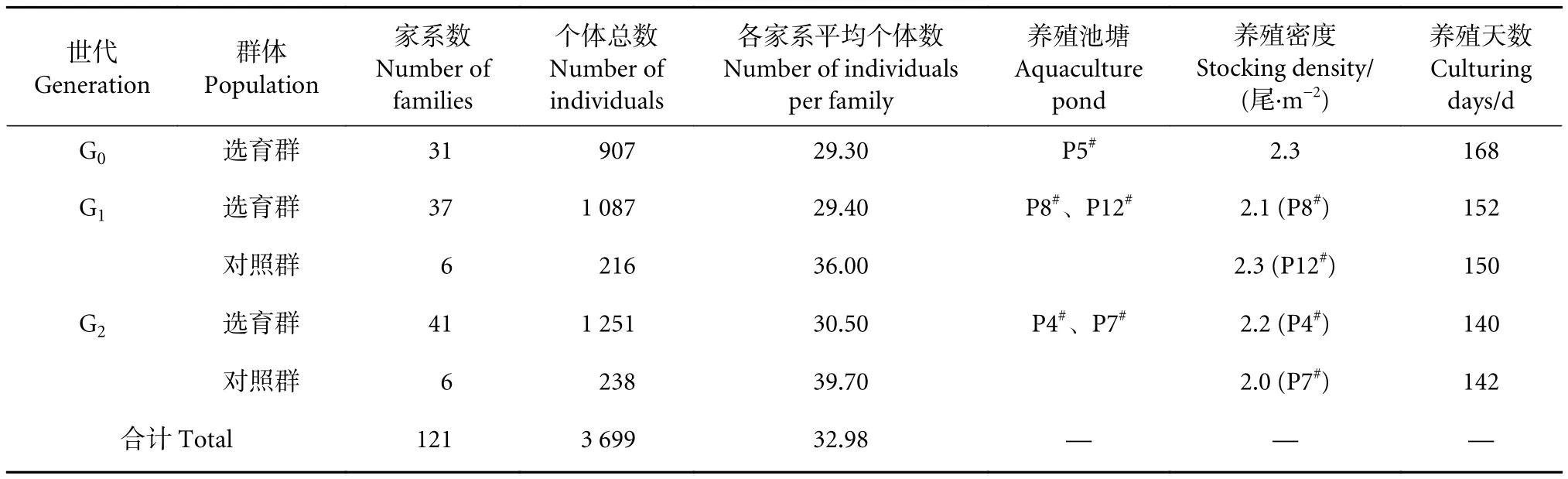
表1 各世代家系构建和培育Table 1 Family construction and breeding for each generation
1.3.3 人工催产与鱼苗孵化
采用促黄体素释放激素类似物 (LRH-A) 进行人工催产,使用计量为雌鱼 10 μg·kg−1、雄鱼5 μg·kg−1;产卵后每个家系选取 10 000 粒以上鱼卵在1 m3的养殖桶中孵化。
1.3.4 鱼苗培育
等仔鱼孵化后,每个家系各随机选取鱼苗2 000尾放入大小为1.0 m×1.0 m×0.8 m的网箱中养殖,待仔鱼长到2~3 g·尾−1时,每个家系随机选取100尾移到大小为2.0 m×3.0 m×1.0 m的网箱中继续养殖。
1.4 标记混合养殖与性状测量
待禾花鲤生长至7~10 g·尾−1时,采用PIT鱼类芯片 (1.35×8 mm) 对其进行标记,每个家系标记50尾,在池塘中混合养殖。其中,G0在1口池塘 (P5#) 中养殖,养殖密度为2.3尾·m−2,养殖时间168 d;G1在2口池塘 (P8#、P12#) 中养殖,养殖密度分别为2.1和2.3尾·m−2,养殖时间分别为152 和 150 d;G2在 2口池塘 (P4#、P7#) 中养殖,养殖密度分别为2.2和2.0尾·m−2,养殖时间分别为140和142 d (表1)。饲料 (质量分数:粗蛋白≥30.0%,粗纤维≥13.0%,粗灰分≤15.0%,钙0.50%~2.00%,总磷≥1.0%,赖氨酸≥1.40%,粗脂肪≥4.0%) 按照实验鱼体质量的5%投喂,每10 d调整一次。当群体平均体质量达到约30 g·尾−1时,捕获其中60%以上的个体进行体质量和体长测定。测量时,将其放入质量浓度为100 mg·L−1的MS-222麻醉剂中快速麻醉,用电子天平测定体质量,精确度为0.1 g。采用游标卡尺测量体长,精确度为0.1 mm。G0、G1和G2世代累计测定家系121个,个体3 699尾。
1.5 数据分析
1.5.1 描述性统计
采用SSPS 22.0软件对各世代的体质量和体长数据描述性统计分析,计算平均值、标准差和变异系数。采用Kolmogorov-Smirnow法进行正态分布检验,拟合回归曲线。
1.5.2 方差分量和遗传力估计
体质量和体长的方差分量和遗传力估计使用ASReml-R软件估计,模型参考Sui等[13]使用的个体动物模型,表达式如下:
![]()
式中:yijklmn表示个体的性状测定值;μ表示总体平均值;Geni表示第i世代的年份效应;Sexj表示性别效应;Pondk表示养殖池塘效应;Geni×Sexj、Sexj× Pondk表示性别与世代、池塘之间的互作效应;Agel( Geni× Sexj× Pondk) 是线性协变量;al是加性遗传效应;fm是家系在混合养殖之前单独饲养时的随机效应;eijklmn是个体随机残差。
1.5.3 表型相关和遗传相关
采用双变量模型估计世代内和世代间的体质量与体长的表型相关和遗传相关。其中,世代内估计时模型中去除世代效应 (Geni),世代间估计时所用模型与式 (1) 相同。
1.5.4 选择反应
G0—G2代选育群的选择反应分别采用目标性状最小二乘值 (方法A) 和基于目标性状育种值 (方法B) 2种方法估计。前者遗传获得为选育群与对照群的性状最小二乘平均值的差值,选择反应用差值占对照群平均最小二乘平均值的百分比表示。后者遗传获得为世代间育种值平均数的差值,选择反应用差值占对照群平均最小二乘值的百分比表示。
每代选择群和对照群性状的最小二乘值估计模型表达式如下:
![]()
式中:Popi表示选择群和对照群的固定效应;其余参数与(1)式相同。
G0—G2代总遗传增益采用下式计算[13]:
![]()
式中:P1为G0—G1代间的选择反应;P2为 G1—G2代间的选择反应。
2 结果
2.1 描述性统计量
全州禾花鲤G0—G2代平均体质量介于33.44~40.88 g,变异系数最低为0.5,最高达0.72;平均体长介于95.01~100.52 mm,变异系数最低为0.18,最高为0.22 (表2);各世代中雌性个体的平均值略大于雄性;其体质量性状较高的变异系数体现了该群体的生长速度具有较高的分化程度,反映了禾花鲤选育群体具有较高的遗传多样性和杂合度。
![]()
表2 体质量和体长的描述性统计量Table 2 Descriptive statistics of body mass and body length
2.2 体质量与体长的正态性检验
分别对各世代体质量和体长指标进行正态分布分析,结果显示各世代数据和整体数据均不符合标准正态分布 (P<0.05)。其中,体质量指标G0、G1和G2代偏度为2.417、1.869和2.297,峰度为13.44、5.58和9.43,总体偏度2.186,峰度9.08,均为右偏分布 (图1-a);体长指标G0、G1和G2代偏度为0.39、0.53和0.58,峰度为0.30、0.35和0.37,总体偏度0.52,峰度0.36,近似正态分布 (图1-b)。在家系培育过程中,少数个体在幼苗期获得的体质量优势在往后养殖中表现出较大的饵料获取优势,因此其肥满度较大,进而在体质量指标方面显著超越正常个体,但体长的增加幅度与体质量增加幅度一致,因此体质量正态分布偏度参数较高 (1.869~2.417),表现为右偏分布 (表3)。
![]()
图1 体质量和体长性状的数据分布Fig. 1 Normal distribution of body mass and body length
![]()
表3 体质量与体长性状的正态性检验Table 3 Normality test of body mass and body length
2.3 方差分量和遗传力
分别利用G0、G1和G2数据估计的体质量遗传力为0.097 (P<0.01)、0.118 (P<0.05) 和0.074,体长遗传力为0.176、0.091和0.099,世代间显示出较大差异,且为低遗传力,其中仅有G0和G1遗传力估计具有显著性 (表4)。可能是利用世代内数据估计遗传力对系谱信息的利用不足,导致加性遗传方差的过低估计和环境方差的过高估计,导致遗传力估计偏低。利用世代间 (G0—G2) 数据估计的体质量遗传力为0.132 (P<0.01),体长遗传力为0.122(P<0.01),在数值上高于利用世代内数据估计的遗传力30%以上,且差异显著 (表4)。表明世代间数据更有利于提高遗传方差的估计准确性。
![]()
表4 方差分量和遗传力Table 4 Variance components and heritability
2.4 体质量与体长遗传相关和表型相关
首先对体质量与体长指标进行相关性分析,结果显示体质量与体长指标极显著相关 (P<0.01),Pearson相关系数为0.91。并以体长为自变量、体质量为因变量进行曲线拟合,获得幂相关函数为y = 0.000 1x2.72(图2)。其次,采用双变量模型估计体质量与体长的遗传相关和表型相关,结果显示世代内估计的遗传相关差异较小 (G0: 0.764, G1: 0.781,G2: 0.757),表型相关具有较大波动 (G0: 0.875, G1:0.837, G2: 0.794)。世代间估计的遗传相关为0.921(P<0.01),表型相关为0.995 (P<0.01,表5),均高于利用世代内数据估计的结果,可能是世代间数据包含更完整的亲缘关系和环境因子,在估计时更好地消除了其对结果的影响。
![]()
表5 遗传相关和表型相关Table 5 Genetic correlation and phenotypic correlation
![]()
图2 体质量和体长的幂相关图Fig. 2 Power correlation plot of body mass and body length
2.5 选择反应
采用2种方法估计选择反应和遗传增益,结果显示基于目标性状育种值估计的体质量性状选择反应为1.89 g (G0—G1)和2.91 g (G1—G2),总选择反应为4.80 g,总遗传增益为12.82%。体长性状选择反应为 1.05 mm (G0—G1)和 1.64 mm (G1—G2),总选择反应为2.69 mm,总遗传增益仅为2.76%(表6);基于目标性状最小二乘值估计的体质量性状选择反应为3.23 g (G0—G1)和2.51 g (G1—G2),总选择反应为5.74 g,总遗传增益为15.45%。体长性状选择反应为4.2 mm (G0—G1)和2.2 mm (G1—G2),总遗传增益为6.60% (表6)。显然,体质量性状获得了较大早期选择反应,而体长性状仅有微弱的选择反应,这可能与亲本选择时采用体质量进行评价有关。
![]()
表6 体质量和体长性状的遗传进展估计Table 6 Estimation of predicted genetic gain for body mass and body length
3 讨论
3.1 体质量和体长性状的表型参数
体质量和体长性状均匀度是养殖鱼类重要的经济性状,较低的体质量离散程度意味着同批次的养殖产品具有较高的一致性,可同批上市,有利于减少劳动强度和降低成本。目前,多数养殖鱼类测定的体质量变异系数介于0.15~0.50[14],其中体质量变异系数较低的品种如罗非鱼 (Oreochromis niloticus) 约为0.18~0.21,经过多世代选育后的优秀品系可低于0.10[15],变异系数较高的品种如鳜(Siniperca chuatsi) 体质量变异系数为0.43~0.55[16],类似的研究结果也出现在牙鲆 (Paralichthys olivaceus)[17]、银大麻哈鱼 (Oncorhynchus kisutch)[18]、瓯江彩鲤[19]等物种中。而体长指标的变异系数较小,如4~6月龄的哲罗鲑 (Hucho taimen) 为0.07~0.09[20],16月龄的草鱼 (Ctenopharyngodon idellus)为0.09[21],总体远低于体质量变异系数。在本研究的养殖条件下,对全州禾花鲤的测定结果表明,养殖142~168日龄的禾花鲤体质量变异系数最低为0.50、最高达0.72,与目前养殖品种相比处于最高水平,从数据组成角度分析,造成变异系数大的直接原因是群体中大个体离群现象突出,数据正态分布偏度值较高 (2.29),类似现象在三角帆蚌 (Hyriopsis cumingii) 的早期养殖阶段也有表现[22]。体长变异系数最低为0.19、最高为0.22,远高于上述牙鲆、瓯江彩鲤等品种。本研究结果表明全州禾花鲤的种群均匀度极低,具有很高的改良潜力。在当前养殖实践中,全州禾花鲤通常采用分阶段过筛的方式获得体质量相对一致的个体出售,无疑增加了劳动力成本的投入;因此,采用科技手段提高该群体均匀度是其种质改良的重要目标之一。
3.2 选育性状的遗传评估
在水产选择育种中,科学可靠的遗传评估对提高选择效率具有重要作用,获得的相关参数是制定配组方案和筛选亲本的重要依据。现阶段,对养殖规模较大品种的选育群体开展遗传评估已极为广泛,主要针对生长、体色、抗病等重要经济性状开展,评估内容包括遗传力、育种值、遗传相关等关键遗传参数[23]。同时,遗传评估结果受多方面因素的影响,如生长阶段、生长环境、遗传参数估计模型等。因此,同品种鱼类各个性状遗传力在不同报道中会出现不同程度的差异。如唐瞻杨等[24]研究不同月龄尼罗罗非鱼数量性状遗传力,3月龄时体质量性状遗传力为0.66,4月龄时为0.17,5月龄时为0.25;罗坤等[25]研究发现,150和410日龄俄罗斯鲟 (Acipenser gueldenstaedtii) 体长的遗传力分别为0.16和0.18,体质量的遗传力分别为0.18和0.09,出现不同生长阶段遗传力差异,但都相对准确地描述了各自选育群体的情况;本研究采用混合线性模型,纳入了选育过程中的年份效应、性别效应、养殖池塘效应、系谱等因素开展体质量、体长性状的遗传力、遗传进展评估,结果表明体质量性状遗传力为低遗传力水平 (h2<0.15),与Hane等[26]对鲤早期体质量性状的遗传力测定结果接近,但低于Wang等[19]估计的鲤体质量性状遗传力。利用双变量模型估计体质量与体长性状的表型相关为0.99,与鳜 (0.96)[16]、俄罗斯鲟 (0.91)[25]等品种的估计值接近。估计的遗传相关为0.92,这与韦信键等[27]对1月龄大黄鱼 (Larimichthys crocea) 的估计结果 (0.83~0.90) 十分接近,类似的结果在团头鲂(Megalobrama amblycephala)[28]、红鳍东方鲀 (Takifugu rubripes)[29]等物种中也有体现,本文获得的遗传参数应在合理区间内;但是,本项目实施过程中每个世代构建的选育家系数量 (31~41) 和对照家系数量 (6) 偏少,选育世代也较少,可能造成的误差依然较大,另外受年份和养殖环境的影响较大,环境方差可能比真实值小,在往后的育种过程中应通过增加家系数量和对照群体数量,优化配组方案,从而提高遗传评估的准确性。
3.3 选育进展
获得选育进展是所有育种项目的最终目标,传统选育方法的革新和分子育种技术的应用使选育进展的获取效率显著提高,根据Gjedrem[30]的总结,在良好的选育方案下水生动物种群的每一代遗传增益约为10%~20%,据报道大盖巨脂鲤 (Colossoma macropomum) 体质量遗传增益可达30.93%[31]。本研究采用的传统家系选育方法,在完成2个世代选育后获得了15.45%或12.82%的体质量遗传增益,而体长性状遗传增益仅为6.60%或2.76%。其中,体质量遗传增益与鲤 (7%)[32]、大菱鲆 (Scophthalmus maximus, 8.70%)[33]、深凹壳型香港牡蛎(Crassostrea hongkongensis, 9.09%)[34]较为接近,但低于罗非鱼平均每代的遗传增益 (13.6%)[35]。体长性状在本次选育中未纳入选育指标,获得的遗传增益为遗传相关带来的间接选择效应。与上述品种相比,本研究获得的遗传增益处于相对较低的水平,也说明本研究采用的选育方法有待进一步改进。此外,本文采用基于目标性状最小二乘值法获得的遗传增益更高,说明在2个世代的选育中现实遗传增益大于预测遗传增益。其原因可能是评估数据中加性遗传效应和环境效应之间存在一定程度的混杂[36],以及家系中缺少半同胞家系系谱信息,从而使加性遗传方差分量被低估[13]。因此,采用基于目标性状最小二乘值法评估的遗传增益更接近真实值。在选育策略和亲本配组方法上,本研究控制近交的同时仅应用传统的家系内选择和1对1配对策略完成了2个世代的选育。目前,有利于遗传评估和提高选育效率的配对方法较多,如在中国对虾(Fenneropenaeus chinensis)[13]上采用最佳遗传贡献理论,在育种核心群长期遗传进展与近交水平的平衡方面优势明显。在亲本配组方法上,可采用更有利于遗传评估的嵌套交配设计,以优化选育过程中的评估准确性。鉴于禾花鲤育种过程中先对家系分池饲养、然后电子标记混合饲养的技术流程,鲁翠云等[37]提出的大规模混合家系选育技术也适用于该品种的选育,应在此后的育种过程中借鉴采用。此外,该群体早期选育遗传增益较低的另一个原因可能在于选育基础群较小,G0选育群仅有31个家系,少于通常采用的选育群家系数量[13,35],导致群体选育潜力未能充分挖掘。前期对全州禾花鲤多样性的研究结果显示其群体遗传多样较高[38],结合其在广泛区域内由农户自繁、自养,而未进行高强度选育推广的实际情况,该群体应拥有较高的选育改良潜力。因此,进一步完善和优化选育方法,选择操作简便、选择强度大的技术路线开展相关工作,结合全州禾花鲤1龄性成熟的特点,有望在短期内获得可观的选育效果。
声明:本站属公益性没有商业目的的网站,上列文章仅供个人学习参考。本站所发布文章为原创的均标注作者或来源,未经授权不得转载,许可转载的请注明出处。本站所载文章除原创外均来源于网络,如有未注明出处或标注错误或侵犯了您的合法权益,请及时联系我们!
欢迎关注本站(可搜索)"养鱼E线"微信公众帐号和微信视频号"养鱼一线"以及头条号"水花鱼@渔人刘文俊"!
