
论文:长江上游5个草鱼群体的遗传多样性
长江上游5个草鱼群体的遗传多样性
翟东东,蔡 金,喻记新,刘红艳,陈元元,熊 飞,段辛斌,刘绍平,陈大庆
(1.江汉大学生命科学学院,湖北省汉江流域特色生物资源保护开发与利用工程技术研究中心,武汉 430056;2.江汉大学,持久性有毒污染物环境与健康危害湖北省重点实验室,武汉 430056;3.中国水产科学研究院长江水产研究所,武汉 430223)
草鱼(Ctenopharyngodonidellus),隶属于鲤科雅罗鱼亚科草鱼属,和青鱼、鲢、鳙合称“四大家鱼”,是我国重要的淡水鱼类资源[1]。然而,由于过度捕捞、水电开发、水体污染等因素的影响,从20世纪80年代起,长江四大家鱼种群数量急剧下降,资源严重衰退[2-3]。长江四大家鱼资源保护和恢复颇受关注,但相关研究主要集中在三峡大坝以下的长江中下游江段[4-6],关于长江上游的研究相对较少。目前,关于长江上游草鱼的研究主要集中在资源量等方面[7-9]。在遗传资源方面,傅建军等[10,11]利用线粒体D-Loop区序列和微卫星标记报道了木洞、万州江段草鱼的遗传多样性和群体结构,廖小林等[12]利用微卫星标记报道了宜宾江段草鱼的遗传多样性。目前,对三峡大坝、向家坝等水电工程建设后长江上游草鱼的遗传多样性及群体结构还缺乏全面的了解。
本研究以长江上游5个不同群体的草鱼为研究对象,通过线粒体Cytb基因研究其遗传多样性、种群分化和历史动态,旨在较全面地了解三峡大坝、向家坝等水电工程开发建设后长江上游草鱼的遗传多样性格局,为长江上游草鱼等家鱼的遗传多样性保护和管理提供科学依据。
1 材料与方法
1.1 样本采集
本研究的草鱼群体样本于2017-2019年采集自长江上游5个不同的江段,分别为邵女坪、巴南、万州、太平溪和合川。邵女坪位于向家坝库区,巴南、万州、太平溪分别位于三峡库区库尾、库中和库首,合川位于长江上游一级支流嘉陵江。邵女坪江段和巴南、万州、太平溪等江段被向家坝阻隔,合川和巴南、万州、太平溪被草街水电站、井口水电站阻隔(图1)。物种鉴定主要参考《中国动物志 硬骨鱼纲 鲤形目(中卷)》[13]。采集的草鱼的标准体长、体重和年龄数据见表1,另外取鱼体背部肌肉保存于95%的酒精中,并存放于-20 ℃,以用于DNA提取。
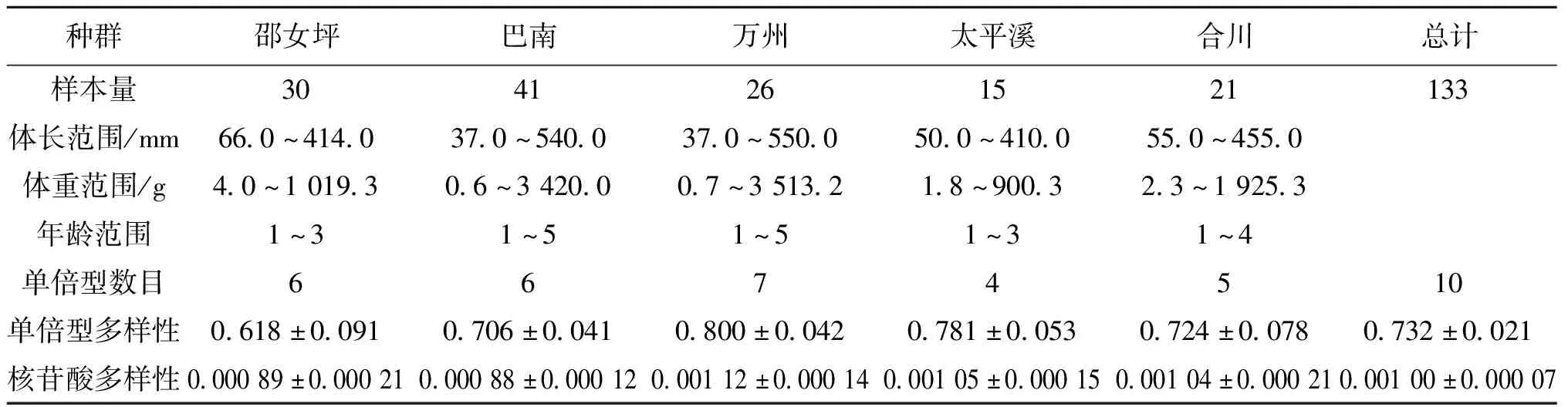
表1 长江上游5个草鱼群体的样本信息和遗传多样性
![]()
图1 长江上游草鱼采样点
1.2 基因组DNA提取以及PCR扩增、测序
取肌肉约50 mg,利用成都福际生物技术有限公司的动物组织基因组DNA提取试剂盒提取基因组DNA。线粒体DNA Cytb片段的扩增引物为L14724 5′-GAC TTG AAA AAC CAC CGT TG-3′(正向)和H15915 5′-CTC CGA TCT CCG GAT TAC AAG AC-3′(反向)[14]。聚合酶链式反应(Polymerase Chain Reaction,PCR)的反应总体积为30 μL:15 μL 2×Taq PCR MasterMix(含Taq DNA聚合酶、dNTPs、MgCl2、反应缓冲液,北京艾德莱生物科技有限公司),模板DNA 3 μL,正反向引物各1 μL,ddH2O 10 μL。PCR扩增程序如下:94 ℃预变性4 min,然后进行35个循环,循环步骤为94 ℃变性45 s,55 ℃退火45 s,72 ℃延伸1 min,最后再72 ℃延伸10 min。PCR产物经琼脂糖电泳验证后,将目的条带准确清晰的样品送至天一辉远生物科技有限公司,完成纯化回收和序列测定。
1.3 数据分析
使用MEGA7对序列进行比对,然后参照测序峰图,对序列进行编辑和校正[15]。运用DnaSP v5.10计算单倍型数目、单倍型多样性(Hd)和核苷酸多样性(Pi)[16]。使用Arlequin version 3.0[17]进行分子方差变异分析(AMOVA)和计算成对遗传分化指数(FST)[18]。通过MEGA7来构建NJ(Neighbor-joining)和UPGMA(Unweighted pair group method using arithmetic average)系统发育树[15]。使用Network 4.6中的中接法(Median-joining)构建单倍型网络图[19]。通过DnaSP v5.10软件进行错配分析来验证群体在历史上是否发生过种群扩张[16]。
2 结果
2.1 序列变异特征
本研究总计获得了133个个体的Cytb全序列,序列长度为1 140 bp,序列中无碱基的插入和缺失。平均碱基组成为A+T的含量为58.3%,G+C的含量为41.7%。在这133条Cytb序列中,共有10个变异位点,占总位点数的0.88%,其中有6个为简约信息位点,另4个为单一变异位点。
2.2 遗传多样性与历史动态
本研究总共获得了10种单倍型(GenBank登录号:MN923194-MN923203),每一个群体的单倍型数目为4到7种。5个群体的单倍型多样性变化范围为0.618到0.800,邵女坪群体最低,万州群体最高,整体单倍型多样性为0.732;核苷酸多样性的变化范围为0.088%到0.112%,巴南群体最低,万州群体最高,整体核苷酸多样性为0.100%(表1)。
邵女坪、巴南、万州、太平溪、合川群体的错配分析图都呈单峰分布,表明这5个草鱼群体在历史上都发生过种群扩张(图2)。
![]()
图2 5个草鱼群体和整个群体的错配分析图
2.3 遗传分化
分子方差变异(AMOVA)分析显示96.96%的变异来自群体内部,仅有3.04%的变异来自群体之间,表明长江上游草鱼群体的遗传变异主要来自群体内部(表2)。群体之间的遗传分化指数FST值如表3,结果显示邵女坪群体和合川群体之间存在显著的遗传分化,其余群体之间都不存在遗传分化。基于群体之间的遗传距离构建的UPGMA系统发育树也表明邵女坪群体与合川群体的遗传距离比其它群体之间的遗传距离要大(图3)。
![]()
图3 基于Cyt b基因序列构建的草鱼群体UPGMA系统发育树
![]()
表2 基于线粒体Cyt b基因的5个草鱼群体的分子变异方差分析
![]()
表3 基于Cyt b基因分析的5个草鱼群体的成对FST值
2.4 系统发育关系分析
NJ系统发育树表明这10种单倍型分成了2支,单倍型网络图结果与NJ系统发育树吻合。通过单倍型网络图可知2个分支都被5个种群所共享,Hap1,Hap2和Hap4是3个最主要的单倍型,都被5个群体所共享,Hap3、Hap6、Hap9、Hap10虽属于私有单倍型,但个体较少,分别只有一个个体,因此整体上没有形成明显的系统地理格局(图4)。
![]()
图4 基于Cyt b基因序列构建的草鱼10种单倍型关系图
3 讨论
3.1 遗传多样性和历史动态
单倍型多样性和核苷酸多样性是评价种群遗传多样性水平高低的两个重要参数,其数值越高,表明群体的遗传多样性越高[20]。Lan和Shi[21]认为当核苷酸多样性数值在0.15%~0.47%之间时,表明群体的遗传多样性较低。本研究5个草鱼群体的整体单倍型多样性为0.732,处于中等水平;核苷酸多样性为0.1%,处于很低水平,整体而言,长江上游草鱼的遗传多样性较低。该结果与历史研究一致,廖小林等[12]利用微卫星标记得到宜宾草鱼群体的期望杂合度和观测杂合度分别为0.648 9、0.574 1,表明长江上游宜宾草鱼群体遗传多样性较低;傅建军等[10]和Zhao等[22]利用线粒体基因序列结果表明长江上游宜宾、木洞、万州、云阳草鱼群体遗传多样性较低。另外,研究者利用RAPD、mtDNA、SSR等分子标记对长江中、下游部分江段、汉江、湘江、鄱阳湖、洞庭湖等草鱼群体的遗传多样性进行了研究,结果也表明草鱼的遗传多样性较低[10,12,22-23]。
长江水系草鱼遗传多样性较低的一个重要原因可能是历史上遭受了瓶颈效应和种群扩张。Grant和Bowen[24]研究结果表明,当Hd≥0.5、Pi<0.5%时,表明群体是受瓶颈效应后迅速扩张导致。本研究中5个草鱼种群的单倍型多样性都较高(Hd≥0.5),而核苷酸多样性较低(Pi<0.5%),说明本研究中的5个草鱼种群是遭受了瓶颈效应之后迅速扩张而来。另外,研究表明错配分析图呈单峰分布,表明群体经历了种群扩张;呈双峰分布,表明群体没有经历过种群扩张[25]。本研究中5个草鱼种群的错配分析图都呈单峰分布,也表明这5个种群发生了种群扩张。历史研究在推测长江中、下游草鱼种群历史动态时,发现长江中、下游草鱼种群也可能遭受了瓶颈效应和种群扩张,如Zhao等[22]利用线粒体ND5、ND6、Cyt b和D-Loop四个基因联合分析草鱼历史动态时,推测石首、瑞昌、邗江江段的草鱼群体都可能遭受了瓶颈效应。此外,近几十年来长江过度捕捞、水电开发过度、鱼类栖息地丧失、水体污染等因素也可能加剧了草鱼遗传多样性的丧失。过度捕捞、栖息地丧失、水体污染等因素致使草鱼的资源量减少而导致草鱼的遗传多样性下降。水电开发则使草鱼群体相互隔离,可能通过减小草鱼群体的有效种群大小而增加遗传漂变来减少群体的遗传多样性[26]。
3.2 遗传分化
本研究结果显示邵女坪群体和合川群体之间存在显著的遗传分化,其余群体之间都不存在遗传分化,这种遗传格局的形成可能和水电站大坝的修建以及人工增殖放流活动有关。研究表明水电站大坝的存在会阻碍坝上、坝下群体的基因交流和增加群体内部的遗传漂变来加大坝上群体与坝下群体的遗传差异[26-28]。本研究中邵女坪群体和合川群体之间产生了显著的遗传分化,可能是邵女坪群体和合川群体被3座水电站大坝分隔,导致它们之间无法产生基因交流而产生遗传分化。如图1所描述,邵女坪位于向家坝库区,即在向家坝之上,合川位于嘉陵江下游,合川到嘉陵江入江口之间有两座水电站,分别为草街水电站和井口水电站,这3座水电站使得邵女坪群体和合川群体的草鱼无法进行基因交流。近些年来,向家坝库区、三峡库区和嘉陵江合川等地都举行了多次草鱼增殖放流活动,而关于放流的苗种、亲本的遗传信息并不明确,因此可能在一定程度上影响草鱼种群的遗传分化格局。Gonzalez等[29]研究结果表明增殖放流对广岛湾不同地点的5个黑鲷野生群体的遗传结构产生了一定程度的影响。
4 结论
长江草鱼群体的遗传多样性水平较低,主要原因可能是群体在历史上遭受了瓶颈效应以及近几十年来长江的过度捕捞、水利工程修建、鱼类栖息地丧失、水体污染等因素导致的种群数量锐减。因此,建议加强对长江草鱼种质资源保护,采取全面禁渔、生境修复、原种增殖放流等措施来促进长江草鱼资源恢复和遗传多样性保护。另外,邵女坪群体和合川群体已经产生了明显的遗传分化,因此在保护工作中,应将他们作为不同的管理单元来保护。
致谢:江汉大学生命科学学院罗辰浩、令狐清清、李雯等学生参与了野外样本采集和部分室内实验,谨致谢意!
声明:本站属公益性没有商业目的的网站,上列文章仅供个人学习参考。本站所发布文章为原创的均标注作者或来源,未经授权不得转载,许可转载的请注明出处。本站所载文章除原创外均来源于网络,如有未注明出处或标注错误或侵犯了您的合法权益,请及时联系我们!
欢迎关注本站(可搜索)"养鱼E线"微信公众帐号和微信视频号"养鱼一线"以及头条号"水花鱼@渔人刘文俊"!
