
鱼类淋巴囊肿病的研究进展
鱼类淋巴囊肿病的研究进展
王锡亮 胡国斌 宋珊珊
(中国海洋大学海洋生命学院 青岛 266003)
【摘 要】鱼类淋巴囊肿病是一种慢性病毒性疾病,可以感染100种以上的温水域和低温水域的海、淡水鱼类,给水产养殖业造成了巨大的经济损失。文章综述了近年来鱼类淋巴囊肿病的研究现状,重点介绍了鱼类淋巴囊肿病的病原、传播途径、发病机理、诊断和防治等方面的研究进展,并对今后鱼类淋巴囊肿病的研究趋势进行了展望。
【关键词】鱼类淋巴囊肿病 淋巴囊肿病毒 发病机理 诊断防治
引言
鱼类淋巴囊肿病(lymphocystis disease)是由淋巴囊肿病毒(lymphocystis disease virus,LCDV)感染引起鱼类的一种典型的皮肤和浅表组织慢性病,它是最早发现的鱼类病毒性疾病。该病的普遍症状是病鱼体表长有瘤状物,严重时内脏组织器官也出现病变。鱼类淋巴囊肿病的较大规模流行最先发生在欧洲的牙鲆养殖场,随后在南美和北美的养鱼场中均暴发过;近年来,开始在亚洲流行。在我国,1992年于养殖的云纹石斑鱼中首次发现了淋巴囊肿病。l997年,山东威海地区的牙鲆养殖场大面积暴发了淋巴囊肿病。该病通常不具有致死性,但是病鱼体质瘦弱、生长迟缓、外表异样而失去商品价值,给水产养殖业带来巨大的经济损失。
鱼类淋巴囊肿病的流行性极广,引起了国内外学者的广泛重视;目前已知至少有42科、125种以上的鱼类可以染上此病,其中海水鱼占30科。当前对鱼类淋巴囊肿病的病原、临床症状、组织病理学变化、诊断等已有多方面的报道,但是发病分子机制尚未明确,疫苗研究也没有获得突破性进展。因此,要想控制和攻克此病,仍需付出巨大的努力。
1鱼类淋巴囊肿病毒
l874年,Lowe首次在鲽形目鱼类中发现淋巴囊肿病,认为该病由原虫引起。l962年,Walker等使用电镜观察到LCDV,指出它就是鱼类淋巴囊肿病的病原体。l965年,Walker将LCDV划分到虹彩病毒科(Iridoviridae)淋巴囊肿病毒属(Lymphocystivirus)。根据国际病毒分类学委员会(ICTV)第八次报告, 虹彩病毒科分为5个属,包括感染脊椎动物的淋巴囊肿病毒属、蛙病毒属(Ranavirus)和肿大细胞病毒属(Megalocytivirus)以及感染无脊椎动物的虹彩病毒属(Iridovirus)和绿虹彩病毒属(Chloriridovirus)。虹彩病毒经过提纯结晶后,在斜射光线照射下呈现蓝紫色的虹彩光泽,故被命名为虹彩病毒。
LCDV是一种大的胞浆型DNA病毒。病毒粒子呈六边形、二十面体对称,直径多在130~260 nm不等,由衣壳、中间脂质层和核心体三部分组成。病毒粒子大小随宿主或所在组织的不同而有差异,Heppell等测定的鱼类淋巴囊肿病毒粒子直径甚至达到380nm。病毒颗粒至少含有33条多肽,分子量大小为l4-220kDa。主要衣壳蛋白(MCP)是病毒颗粒的重要结构成分,约占多肽的40%-45%。MCP基因是水生虹彩病毒进化上比较保守的基因,被视为虹彩病毒分子进化的标记。
LCDV基因组大小约l02-186 kb,为一条具有环状变换和末端冗余的线性双链DNA分子,位于核心体中,其中的胞嘧啶5’端高度甲基化。目前,根据MCP基因保守序列将LCDV分为3种基因型:基因型I、基因型Ⅱ和基因型Ⅲ。徐洪涛等从我国养殖牙鲆中分离出的淋巴囊肿病毒中国株(LCDV-C)属于基因型Ⅱ,它不同于欧洲分离的基因型I LCDV-1病毒株,两者MCP的同源性为87.6%。LCDV-1是虹彩病毒中最早测出全长序列的病毒株,其基因组全长为102653bp,包含195个开放阅读框(ORF),编码的多肽分子大小为40-1199kDa。2004年,LVDV-C基因组全序列也被测出,大小为l86250bp,含有240个ORF。比较后发现,两者的编码能力和基因排列顺序都有较大的差异。2007年,Eaton对虹彩病毒科中序列已知的12种病毒基因组进行了分析,并鉴定出26个核心基因,经过重新注释后的LCDV1与LVDV-C基因组序列分别含有1O8和178个ORFs。
![]()
2 传染途径
淋巴囊肿病毒经由水体或直接皮肤接触,通过呼吸器官、消化道和皮肤伤口在同种或不同种鱼之间进行水平传播。绳秀珍预测病毒至少可以通过体表伤口、鳃、消化道上皮等三条途径进入鱼体内。曲径等通过对牙鲆进行皮肤划伤感染实验证实病毒主要以体表感染的方式进行传播。
迄今为止,有关淋巴囊肿病毒在鱼体内的传播途径的研究资料比较缺乏。Colorni认为,淋巴囊肿细胞破裂后会释放病毒粒子到邻近组织中,然后进入血液循环系统。孙修勤等用原位杂交的方法在牙鲆的胃、性腺、肝脏和肌肉组织中检测到病毒,推断病毒经口进入消化道、再进入血液进行传播。刘允坤等用PCR方法检测发现,人工感染淋巴囊肿病毒的牙鲆的血液和脾脏组织中病毒浓度最大,其次是肠、鳃、胃和肝,由于脾、肾脏是鱼类的造血器官,因此推测病毒能很快地随着血液循环扩敖到性腺,进行垂直传播。
通常病毒在细胞间的传播方式有三种:
(1)细胞外传播:受染细胞释放的病毒再去侵染其他细胞;
(2)细胞间传播:病毒感染细胞后并不释放而是通过细胞阔桥传给相邻细胞;
(3)细胞分裂传播:病毒随着细胞的繁殖从亲代传染到子代。
曾晓华通过斑点免疫吸附分析发现,出现明显病变的细胞培养物上清中的病毒粒子极少,表明病毒传播可能以细胞间传播为主,随着病变程度加深,细胞崩解,推测病毒有可能存在细胞外传播方式。吕宏旭等认为淋巴囊肿病毒可通过受体介导和吞噬作用两种细胞内吞方式进入胞内;此外,还通过膜融合进人胞内,这是胞膜病毒最常采用的方式。
当淋巴囊肿病毒侵入鱼体后,首先在受染的表皮细胞之内形成淋巴囊肿细胞。包涵体是病毒在细胞中的增殖场所,病毒在其中装配完毕后进入细胞质,其后细胞质开始衰退,包涵体也消失,在胞浆内可见大量的散在病毒颗粒,直至细胞破裂,病毒颗粒散布到水中,继而感染体表受损的鱼体。集约化养殖在引入新苗种或者水体中含病毒时易感染淋巴囊肿病,然而,由于操作、捕捞、交配、寄生物感染以及鱼体之间的攻击行为等原因导致的皮肤创伤都加剧了病毒在鱼群中的传播;提高水温和养殖密度也能加剧病毒的传播。Kitamura等研究表明,夏天水温高时发病率高,冬天发病率较低。然而,曲径等的调查发现,牙鲆淋巴囊肿病一年四季均可发生,低水温期病情似有加重。张永嘉等报道,在我国广东海区养殖鱼类中,该病的流行季节是10月份至翌年5月的低水温期(10-25℃),苗种发病时死亡率达30%。这与以往认为此病多发生在高水温季节的观点不同,这些差异可能与养殖海区的环境有关。由此可见,淋巴囊肿病的发生与水温没有密切的关系。
![]()
3发病机制
目前,鱼类淋巴囊肿病的发病机制尚不明确 已知鱼类虹彩病毒主要通过诱导细胞凋亡和裂解细胞两种方式损伤受染细胞。淋巴囊肿病毒感染培养细胞后,出现明显的细胞病变效应(CPE),主要表现在细胞固缩,呈牵丝状,然后细胞聚融合坏死,脱落并形成空斑,最终随着空斑逐渐变多并扩大,整个细胞单层被破坏脱落。通过对细胞核形态、DNA片断化、天冬氨酸酶活性等研究,证明LCDV诱导宿主细胞调亡;凋亡的发生基本上不影响子代病毒粒子的复制。根据大菱鲆淋巴囊肿病毒SDS-PAGE图谱分析,来自鱼体囊肿细胞的LCDV与来自细胞系扩增的LCDV有明显差异,后者缺少14,25和45kDa的多肽,推测此三种多肽可能与病毒的致病性相关。胸苷合成酶(Thymidylate Synthase,TS)参与细胞周期调节和肿瘤的形成。病毒感染鱼体后,该酶存在于宿主的细胞质中,并促使细胞进入S期和G2/M期。赵哲在LVDV-C中发现0RF011L含有完整的TS序列,这是第一个发现的脊椎动物虹彩病毒的TS基因,有助于认识和阐明淋巴囊肿病毒致病的分子机理。
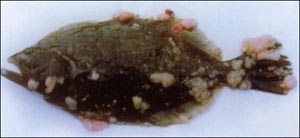
淋巴囊肿病毒偏好感染真皮结缔组织中成纤维细胞和成骨细胞。病毒感染使细胞扩大、增肥,不再进行有丝分裂,从而形成球形的肥大细胞,此时细胞直径可达0.3-2mm;同时细胞核发生退行性变化、凝结或破碎,核仁扭曲并模糊;胞浆发生变化,在细胞质边缘散有嗜碱性的、大小不一的包涵体;在淋巴囊肿周围有一层厚而透明的硫酸粘多糖性质的包膜;在染病早期,包膜有轻微炎症,感染后期淋巴囊肿破裂,诱发更激烈的炎症反应。淋巴囊肿是最早在病鱼体表出现的症状,它们通常生长在皮肤、吻端、眼眶周围、鳍和尾等部位,严重时遍及全身甚至内脏组织器官,如腹膜、心包、咽、肠壁、卵巢、脾、肝等,其颜色从白色到微红色不等,取决于感染的部位和血管的丰富度。
淋巴囊肿细胞破裂后,病毒粒子释放出来,侵染邻近组织并进入血液循环系统,引起全身性感染。囊肿破裂后还易形成开放式溃疡面,导致寄生虫、细菌等其他病原体的继发感染。曲凌云等在患病牙鲆的心肌、脾细胞、肠上皮细胞、鳃细胞和头肾等组织中均观察到了直径略有不同的病毒颗粒,鳃细胞中的病毒数量仅次于体表。这些来自不同组织的病毒粒子的毒力是否相同还有待进一步验证。Colorni等认为,在系统性感染早期,脾细胞起主要免疫作用,但是当宿主的免疫力下降时,脾细胞的吞噬能力降低,导致其他内脏器官的相继感染。Mareogliese等发现,LCDV促进细胞吞噬能力,但不能明显促进非特异性免疫细胞的增生。并推测病毒以整合或非整合形式存在于易感细胞中而干扰细胞的核酸代谢,导致细胞产生不同程度的病变。
患病个体生长缓慢,体形消瘦,一是因为淋巴囊肿瘤状物对病鱼的呼吸与运动造成了障碍而影响了摄食,二是因为随着病毒侵入内脏器官,尤其是心脏与肾脏功能的降低而导致体质恶化。张永嘉认为造成病鱼死亡的主要原因是肾功能衰竭而导致的排泄系统功能障碍。常藕琴通过对军曹鱼淋巴囊肿病的研究认为,病毒引起鳃、心脏、肾脏、脾脏、肝脏、肠道和脑等器官病变而致多器官功能衰竭,其中心肌纤维和肾小管上皮细胞受损是造成病鱼死亡的主要原因。
到目前为止,关于病鱼对再次感染是否具有保护性的看法还不统一。Nishda发现,患病牙鲆自愈后其抗体水平比发病鱼体和健康体高出不少。Yoshimizu等证实LCDV灭活疫苗对牙鲆有保护作用。1wamoto等的研究表明,用HINAE细胞系扩增的LCDV对牙鲆无感染性,而对鱼体有保护作用。Chinchar根据LCDV染病鱼体的体表损伤特性和病发部位的免疫应答特点。推测在阻止病毒感染的过程中,免疫系统的作用是不明显的。一些研究还表明,环境因子的变化似乎可以刺澈淋巴囊肿病的发生,这些因素包括养殖密度、低盐度、季节、水温和环境污染等
4 疾病的检测和诊断
通常淋巴囊肿组织的病理学特征可作为疾病诊断的可靠依据,但是鱼类淋巴囊肿病是一种慢性疾病,从感染病毒到出现组织学上的可察病症需要很长的时间。此外,由于病毒独特的生物学特征和鱼类的水生生长环境使该病的治疗和预防更加困难。因此如何提高诊断方法的敏感度,并采取相应措施防止疾病的流行,是生产中减少损失的最有效的途径。目前常用的诊断方法可归纳为以下几类:
4.1电镜观察
通过电子显微镜可以观察细胞的病理结构、病毒形态大小和类型以及了解病毒的感染、复制和形态发生等特征。l962年。wa1ker通过电镜观察确定鱼类淋巴囊肿病的病原体为淋巴囊肿病毒。徐洪涛通过电镜观察到发病牙鲆病变组织中的球状淋巴囊肿病毒。曲凌云、绳袖珍等使用电镜研究了淋巴囊肿病毒在患病牙鲆中的分布。常藕琴在患病军曹鱼的囊肿细胞质中观察到有大量二十面体的病毒粒子,大多数病毒粒子的核髓和衣壳清晰,结构完整,少量病毒粒子正在装配,有极少致病毒只有不完整的衣壳。
4.2 细胞培养技术
细胞培养技术是病毒学研究的基础,也是病毒病诊断的经典方法之一。寄生于培养细胞中的淋巴囊肿病毒在23℃的条件下可存活数月。1966年,Wolf等利用棘臀鱼(centrarchid)细胞系增殖LCDV获得成功。随后的研究发现,LCDV可在蓝鳃鱼幼鱼细胞系BF-2(bluegill fry-2)、金头鲷鲭细胞系SAF-1 (gilthead seabream fin-1)、亚洲鲈鱼幼鱼细胞系SF(Asian seabass fry)、日本牙鲆胚胎细胞系HINAE (Japanese flounder embryo)、草鱼肾细胞系GCK (grass carp kidney)和草鱼卵巢细胞系GCO(grass carp ovary)等细胞内复制并缓慢引起细胞病变。孙修勤、吕宏旭等分别利用牙鲆鳃细胞系进行了养殖牙鲆淋巴囊肿病毒的分离和增殖。虽然多种细胞系是淋巴囊肿病毒感染的许可细胞,但是淋巴囊肿病毒对它们的感染能力并不相同。LCDV接种于GCO中,只需一天就能观察到典型的细胞病变效应,FG需要6天,SAF-1至少需要l0-15天,BF-2则需要21天才可见典型的细胞病变效应。
4.3免疫检测
免疫检测是基于抗原抗体反应的一种检测方法。当前,许多免疫技术已经应用于LCDV的检测。1998年,日本学者建立了淋巴囊肿病的ELISA诊断法。Garc i a-Rosado等建立了血清学检测方法。比较间接荧光测定、细胞计量术和斑点印迹免疫酶技术对SAF-l细胞中LCDV抗原的检测效果发现,间接荧光溯定法快捷敏感,细胞计量术有漏检情况发生,斑点印迹免疫酶检测则易产生假阳性,因此认为间接荧光确定最适合于SAF-1细胞的LCDV抗原检测。特异抗血清的使用提高了血清学检测方法的敏感性。Cano等利用特异抗血清通过免疫印记技术对带有瘤状病灶的病鱼以及携带病毒但无症状的个体进行了检测。单克隆抗体敏感、易获得,对疾病的诊断具有较高的特异性,同时容易做到诊断程序的标准化。绳袖珍利用抗LCDV的单克隆抗体结合免疫荧光技术对感染病毒的许氏平壹由进行检测,并在淋巴囊肿细胞的细胞质中、胃上皮、鳃片以及肌纤维下表皮检测到特异荧光信号。程顺风等通过抗LCDV的小鼠单克隆抗体,结合ELISA诊断、间接免疫荧光法、蛋白质印迹、免疫电子显徽技术,对LCDV进行检测。发现具有特异反应的LCDV蛋白,井检测到单克隆抗体作用于病毒粒子外表面。
4.4分子诊断
核酸杂交技术是继免疫诊断技术之后产生的第三代诊断技术。孙修勤采用核酸探针杂交方法可有效地对LCDV进行定性和定位诊断,并提出LCDV是通过血液在牙鲆体内传播,同时可能存在垂直和水平两种传播方式。
PCR技术具有敏感、特异、快速和简便等优点,目前已成为检测虹彩病毒的主要技术手段。针对虹彩病毒的MCP、DNA聚合酶(DdDP)、DNA转甲基酶(DMet)、ATP酶(ATPase)、核苷酸还原酶(RBRD)基因以及其他ORFs或限制性酶切核酸片断序列,建立了相应的PCR检测技术,并成功进行了鱼类细胞淋巴囊肿病毒的检测。其中所用的引物大部分都是根据虹彩病毒MCP基因的高度保守区域设计的。此外,Kitamurat利用淋巴囊肿病毒MCP基因的高变区和保守区,建立了多重PCR技术,用于淋巴囊肿病毒不同地理株基因型的鉴定。Cano等利用PCR技术结合狭缝印记杂交技术进一步提高了病毒检测的敏感性,所用的病毒DNA量仅为2.5ng。定量PCR技术被用于确定淋巴囊肿病毒滴度和发病率之间的关系。咎金东通过竞争性定量PCR对牙鲆组织中的LVDV-C进行定量,发现长有明显肿瘤的牙鲆外周血中LVDV-C的平均含量是无瘤染病牙鲆的100倍;当少量病毒感染时,牙鲆并不出现病症。
5预防与治疗
淋巴囊肿病是一种慢性病毒病,感染率达30%,发病不严重的鱼可以自行痊愈,有的病鱼的囊肿可以自行脱落,有的在口唇部影响进食的病鱼则需外科手术摘除。经研究发现,22-25℃高温水结合过氧化氢药浴可使病鱼体表囊肿消失。使用中草药制剂对淋巴囊肿病毒进行预防和治疗,治愈率达80%,用于预防发病率低于10%的鱼群。Yoshimizu等研制出LCDV灭活疫苗,并证实其对牙鲆有保护作用,但是该制备方法不适合规模化生产。核酸疫苗与传统的抗原疫苗相比,可诱导机体产生广泛的体液免疫和细胞免疫,同时具有生产成本低以及保存上的许多优点。目前,已经将淋巴囊肿病毒主要衣壳蛋白基因片段分另别进行了原核和真核表达,并且成功诱导小鼠产生特异体液免疫应答。孙修勤等构建的真核重组质粒可以在牙鲆胚胎细胞FEC中表达。郑风荣等将阳性重组质粒经肌肉接种牙鲆,验证了该疫苗良好的免疫效果和较高的保护率。
迄今还缺乏有效的治疗淋巴囊肿病的措施,因此疾病的预防非常关键。预防养殖群体发生疫病在集约化养殖模式中至关重要,通常采取以下方式:(1)把好苗种关,对亲鱼、苗种严格检疫。发现带毒者,截断疫源、销毁病死鱼;(2)对病鱼实施隔离养殖,并用过氧化氢溶液浸洗,提高养殖水温,同时注意预防并发感染;(3)把好水质关,提倡流水养鱼,加大换水量,过滤消毒水源;(4)把好饵料关,不用鲜活饵料、小杂鱼等,杜绝病原从饵料带入;(5)降低养殖密度以减少发病机会;(6)增加营养,提高鱼类机体自身免疫能力。
6 鱼类淋巴囊肿病的研究展望
自1962年Walker目首次鉴定鱼类淋巴囊肿病毒以来,学者们围绕鱼类淋巴囊肿的细胞发生学和病原分子生物学开展了研究。近年来,在病毒基因组序列测定和注释、发展诊断技术、研制疫苗等方面取得了一定的进展,但还有许多方面的研究有待进一步深入,如病毒装配过程、免疫逃逸机制、宿主感染特异性和特效疫苗研制等方面。
淋巴囊肿病毒在鱼体内潜伏期长,一旦出现病症来不急医治就传播开来,因此如何提高诊断方法的敏感度和有效性对于疾病流行的控制具有重要的意义。应用分子生物学和免疫学技术可以达到早期和快速诊断的目的,从而指导病毒病的防治。快速诊断试剂盒的研制是今后加强疫病防控的手段之一。由于病毒存在于活体细胞中,要求药物具有极高的选择性,因此药物开发难度大;并且长期施药不仅导致水体的污染,也会造成诸多耐药性病原体的产生,使进一步的防治工作变得复杂。核酸疫苗的出现开辟了疫苗学的新领域,但还存在着免疫效果不稳定和安全性等问题。因此,研制安全、稳定、特效的疫苗仍是今后的努力方向。
随着分子生物学技术的发展,淋巴囊肿病毒LCDV-1、LCDV-C基因组全序列的测定为研究工作带来新的生机。利用世界各大数据库(GenBank/EMBL/DDBS)中海量的生物信息资源,并结合各种实验手段,将大大加速了解淋巴囊肿病毒的基因组成及功能,使阐明LCDV感染的分子机制和获得LCDV与宿主细胞相互作用的关键因素成为可能。90年代兴起的人类基因组计划(GHP),积极推动了基因克隆技术的发展,相继出现了数种基于消减杂交策略的基因克隆方法。张义兵等以紫外线灭活的草鱼出血病病毒(GCHV)诱导细胞,构建鱼类细胞抗病毒基因差减cDNA文库,并从文库中分离鉴定多种抗病毒和/或免疫相关基因。因此,可以利用抗病基因进行抗病育种,这也是鱼类疾病防预的一种有效途径。
(参考文献略)
声明:本站属公益性没有商业目的的网站,上列文章仅供个人学习参考。本站所发布文章为原创的均标注作者或来源,未经授权不得转载,许可转载的请注明出处。本站所载文章除原创外均来源于网络,如有未注明出处或标注错误或侵犯了您的合法权益,请及时联系我们!
欢迎关注本站(可搜索)"养鱼E线"微信公众帐号和微信视频号"养鱼一线"以及头条号"水花鱼@渔人刘文俊"!
