
背鳍缺失对胭脂鱼幼鱼游泳能力与反捕食能力的影响
背鳍缺失对胭脂鱼幼鱼游泳能力与反捕食能力的影响
周龙艳,付世建,付 成,凌 鸿,李秀明
(重庆师范大学进化生理与行为学实验室,重庆市动物生物学重点实验室,重庆401331)
游泳运动是鱼类逃逸、捕食和繁殖等行为实现的主要方式,为鱼类的生存、生长和繁衍提供基本保证[1]。鱼类游泳行为复杂多变,且存在显著的种间和种内分化。因此,需要选择合适的指标对鱼类的游泳能力进行客观评价。一直以来,临界游泳速度(critical swimming speed,Ucrit)和最大匀加速游泳速度(constant acceleration test speed,Ucat)被研究者广泛应用于鱼类游泳能力的评价[2]。Ucrit作为鱼类有氧运动能力的重要评价指标[3],主要受到最大有氧代谢能力(maximum metabolic rate, MMR)的限制。而Ucat作为鱼类无氧运动能力的重要评价指标[3]。二者对于鱼类在自然环境条件下逃避捕食、穿越激流以及寻找适宜生境等活动密切相关,具有重要的生态学意义。另外,最近的研究发现尽管运动时间短(不足1 s)且移动距离有限,快速启动逃匿过程(fast-start response)的运动性能对鱼类有效捕获猎物和逃避敌害至关重要[4]。因此,该过程的最大速度、最大加速度、旋转角度以及旋转率(后二者为灵活性的指标)可能与鱼类的生存适合度紧密关联[5]。
鱼鳍是鱼类游泳的重要器官之一,主要包括胸鳍、腹鳍、臀鳍、背鳍以及尾鳍。不同鱼鳍在鱼类游泳过程中发挥着截然不同的作用。作为游泳运动的重要部位,尾鳍与鱼类运动能力的关系一直受到研究者们的广泛关注[6-7]。而背鳍对鱼类游泳能力的影响却鲜有涉及。胭脂鱼(Myxocyprinusasiaticus)属鲤形目(Cypriniformes)胭脂鱼科或称亚口鱼科(Catostomidae),该科鱼类全世界约有 13 属70 种,绝大多数种类分布于北美洲,但是胭脂鱼在我国为特有种,分布于我国的长江和闽江流域[8]。因此,胭脂鱼在鱼类系统分类和动物地理学研究上有着极为重要的科学价值。相关的调查报道指出,目前胭脂鱼在闽江已近绝迹,长江种群的资源也日趋枯竭,目前已经被列为国家二类水生野生保护动物[9]。不同生长阶段的胭脂鱼体形差异明显。幼鱼阶段体型奇特,体较高,呈三角形,背鳍宽展而高大,有“一帆风顺”的寓意[10-11]。本研究主要探讨背鳍在胭脂鱼幼鱼游泳运动和反捕食过程中的作用,为其资源保护提供重要的参考资料。
1 材料与方法
1.1 实验鱼的来源与驯养
胭脂鱼幼鱼(Myxocyprinusasiaticus)于2017年11月购自重庆市北碚区,体质量(2.30±0.12) g, 体长(4.59±0.09) cm,共124尾,在实验室自净化循环控温水槽中驯养21 d,曝气24 h后的自来水作为实验用水。驯养期间每天10: 00采用商业颗粒饲料饱足投喂一次,30 min后用虹吸管排除残饵与粪便;水槽日换水量约为水体(200 L)的10%,水温控制在(25±1)℃,并用充氧泵供氧。驯养结束后,挑选身体健康、体重、体长接近的实验鱼作为实验对象。
捕食者乌鳢(Ophiocephalusargus)购于重庆市永川区渔场。购回的乌鳢放入(25±1)℃的水槽中驯养14 d,期间用曝气24 h后的自来水驯养。
1.2 实验方案
1.2.1 实验设计
剪鳍处理:将实验鱼轻度麻醉(3-氨基苯甲酸乙酯甲基磺酸盐, MS-222,50 mg/L)后用剪刀进行剪背鳍处理(图 1)。切除背鳍处理后的实验鱼放回水体恢复2 d[5]。
游泳能力测定:选取实验鱼60尾用于3种游泳能力的测定,每种游泳能力取20尾测定,其中对照组10尾背鳍缺失组10尾。
捕食死亡率:实验设4个重复,每个重复选取实验鱼16尾用于捕食死亡率的测定,其中8尾进行背鳍切除,另外8尾作为对照组。
1.2.2Ucrit的测定
采用实验室自行研制的游泳代谢测定仪测定Ucrit[12]。具体操作为首先将单尾实验鱼放入测定仪中适应2 h,适应过程的水流速度为4. 0 cm/s(约1 倍体长/秒,Body Length,1 BL/s);随后以4 cm/s的速度增量提高水流速度至 8、12、16和20 cm/s等,各水流速度下的持续游泳历时均为20 min[1]。重复上述操作直至实验鱼表现出力竭状态即停止实验。力竭的判断标准为实验鱼被水流冲至游泳管尾端筛板,且身体呈卷曲状态的时间不低于20 s[13]。计算公式为:
Ucrit=[V+(t/ΔT)ΔV]
(1)
式中:V为顺利完成设定时间的最大游泳速度(力竭前一档速度);t为未能完成当前速度梯度下设定时间的实际持续游泳时间(t<20 min);ΔT为各速度梯度下的持续时间(即20 min);ΔV为各速度梯度的速度增量(即 4 cm/s)。
运动代谢率在测定Ucrit的过程中进行测定。具体操作为每2 min测定 1 次游泳代谢仪封闭水体的溶氧值,以溶氧值随时间变化斜率的绝对值和水体的体积计算每尾实验鱼的运动过程的耗氧率(MO2)
作为代谢率的指标。实验结束后将实验鱼取出,对游泳代谢仪重新密闭并进行 20 min 的耗氧率测定,用以消除细菌耗氧对实验鱼MO2的影响[13],计算公式为:
MO2=(St-S0)×3.45× 60/(W× 1 000)
(2)
式中MO2(mgO2/(kg·h))代表耗氧率,St为每档期间(20 min)溶氧值随时间变化的斜率,S0为细菌耗氧的溶氧值变化斜率,3.45 为游泳代谢仪体积(L), 60 为 60 min。W表示实验鱼的体重(g)。
1.2.3Ucat的测定
Ucat测定的设备与Ucrit基本相同,主要改进之处为调节流速的方式改变为通过智能化调节输出电源的频率而改变电机的转速。具体操作如下:将实验鱼转入游泳代谢测定仪中驯化适应2 h,流速为4 cm/s。驯化结束之后,将测定仪中的水流速度以恒定的加速度(10 cm/s2)持续均匀增加水流速度,实验鱼力竭时的水流速度即为Ucat [14]。
1.2.4 快速启动游泳能力的测定
实验前将直径 1 mm 的白色圆形纸片固定于实验鱼背部中央皮肤表层,作为实验鱼的质心标记点[15],以便进行图像分析和参数计算。拍摄整个快速启动过程并计算快速启动指标。实验结束后,用图像处理软件(ACD see v 8. 0)对拍摄的图像进行处理并采用软件(tpsdig2)进行数据采集和分析。通过计算,分别求得实验鱼质心的最大线加速度(Amax)、速度(Vmax)以及旋转率和旋转角度,参数计算详见相关参考文献[4,16]。
1.2.5 捕食死亡率的测定
将对照组和背鳍缺失组各8尾实验鱼一同转移至提前放置有2尾捕食者的捕食测定水槽中(长1.5 m×宽0.8 m×高0.5 m)。在实验过程中,每3 h观察一次捕食情况,当一半的胭脂鱼(即8尾)被捕食时,停止实验。设置4个重复。捕食死亡率的计算为被捕食的对照组或背鳍缺失组个体占总被捕食个体的百分比[17]。
1.2 统计分析
实验数据采用SPSS 17进行统计分析。统计值均以平均值±标准误(Mean±SE)表示,显著水平为P<0.05。对照组和背鳍缺失组之间各参数的差异使用t检验进行统计分析。背鳍缺失和游泳速度对运动代谢率(MO2)的影响采用重复度量的双因素方差分析。
2 结果
2.1 背鳍缺失对胭脂鱼幼鱼Ucrit及其代谢率、Ucat和快速启动能力相关参数的影响
对照组与背鳍缺失组的Ucrit及各代谢参数均无显著性差异(表 1)。随着水流速度的增加,对照组和背鳍缺失组的MO2均显著上升,但对照组和背鳍缺失组在同一游泳速度下的MO2均无显著差异(图2),暗示其组间游泳效率无显著差异。对照组与背鳍缺失组胭脂鱼幼鱼的Ucat没有显著性差异,分别为(53.12±3.02)和(52.92±1.52) cm/s(表 1)。对照组和背鳍缺失组的Vmax分别为(1.12±0.08),(1.11±0.05) mm/ms;Amax分别为(0.22±0.02),(0.19±0.01) mm/ms2;旋转角度分别为(54.91±3.3)(°),(59.66±7.33)(°);旋转率分别为(1 390.10±171.03,(1 762.51±266.83)(°)/s。对照组与背鳍缺失组快速启动各个参数均无显著差异(表 1)。
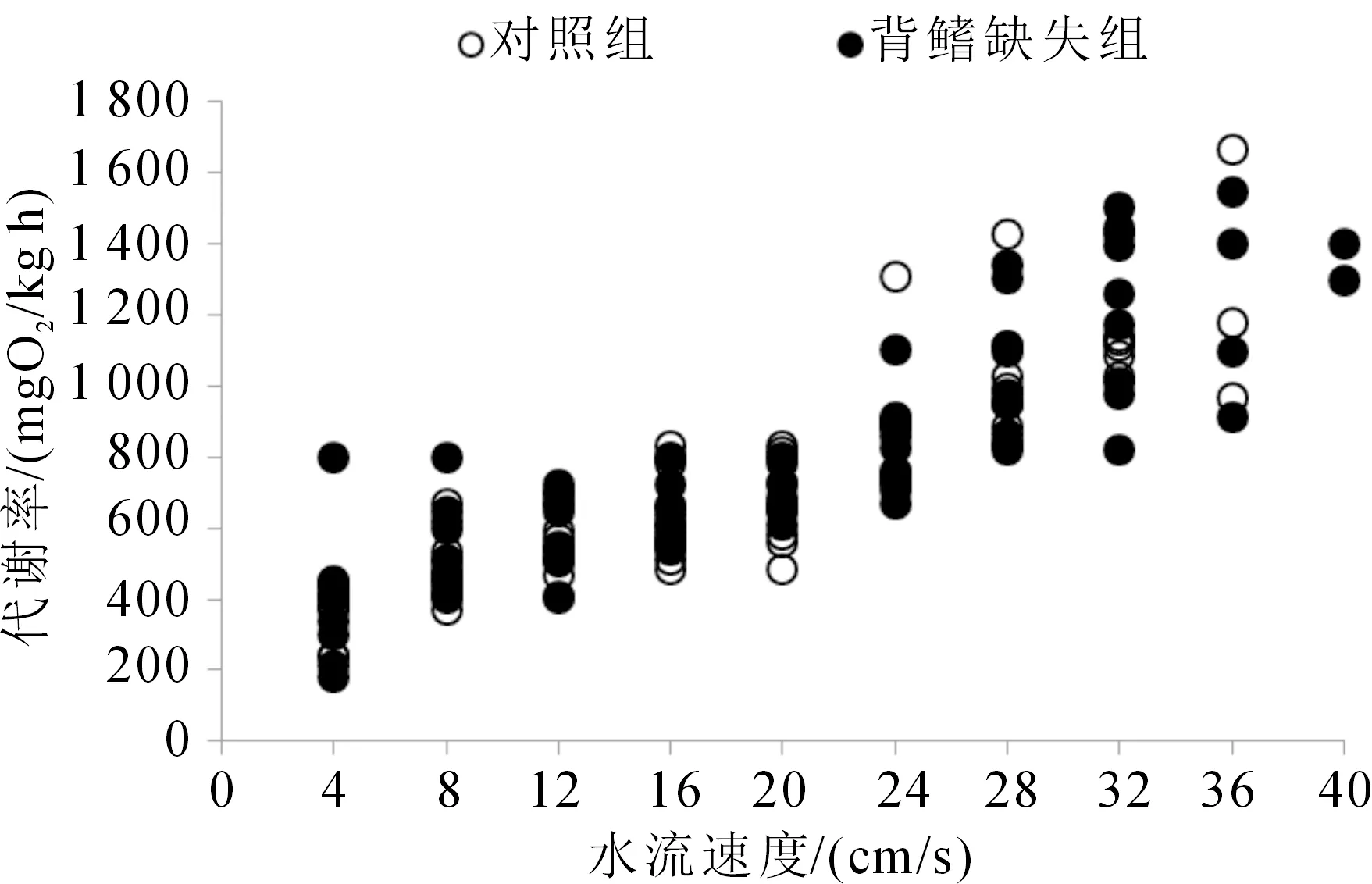
图2 背鳍缺失对胭脂鱼幼鱼游泳过程中耗氧率的影响
2.2 背鳍缺失对胭脂鱼幼鱼在捕食者存在条件下存活率的影响
从图3可以看出,对照组胭脂鱼幼鱼在捕食者存在条件下的存活率为62.5%,而背鳍缺失组胭脂鱼幼鱼在捕食者存在条件下的存活率为40.6%。背鳍缺失对胭脂鱼幼鱼在捕食者存在条件下存活率具有显著影响,背鳍缺失组胭脂鱼幼鱼的存活率显著低于对照组(P<0.05)。
![]()
图3 背鳍缺失对胭脂鱼幼鱼反捕食能力的影响
3 讨论
鱼鳍在鱼类游泳过程中起着重要的作用,不同类型、大小和形态的鱼鳍在鱼类运动过程中的作用不尽相同[18]。尾鳍作为运动系统的重要器官, 在游泳过程中具有推进及导向作用,因此大部分研究主要关注鱼类尾鳍与运动能力的关系[5-7]。尾鳍的缺失导致鳊(Parabramispekinensis)、金鱼(Carassiusauratus)、中华倒刺鲃(Spinibarbussinensis)和鲤(Cyprinuscarpio)幼鱼的Ucrit分别下降40%、35%、49% 和 32%[7,19-20]。由于胭脂鱼幼鱼具有特殊的背鳍形态,可能对其游泳能力产生一定的影响。不过,相关研究还未见报道。本研究通过比较对照组和背鳍缺失组胭脂鱼幼鱼之间Ucrit发现,背鳍缺失对胭脂鱼幼鱼持续游泳速度没有显著的影响(表 1)。这与唐鱼(Tanichthysalbonubes)切除背鳍后的研究结果相似[20]。此外,由于鱼类的游泳能力与其能量代谢能力密切相关,一些研究者也对鳍缺失后鱼体耗氧率进行了研究[6,19]。不过,以往的研究发现尾鳍缺失并没有对红大马哈鱼(Oncorhnchusnerka)和鳊的MMR产生显著影响[6,19]。本研究发现,背鳍缺失处理后胭脂鱼幼鱼的RMR、MMR和MS等代谢参数也都没有产生显著性变化(表 1)。由此表明,背鳍的缺失并没有对胭脂鱼幼鱼游泳代谢能力产生影响(图 1),这可能是其游泳能力没有产生显著变化的原因之一。总体而言,鱼类背鳍在游泳过程的作用远小于尾鳍,胭脂鱼进化过程中形成的独特的背鳍形态及其生态学作用值得进一步研究。
鱼类生存能力与其游泳能力密切相关,特别是鱼类非稳定游泳能力(通常用Ucat和快速启动过程中的Vmax、Amax、旋转角度以及旋转率等指标表示)在鱼类捕食和避敌过程中发挥着重要的作用[3-5]。研究发现,尾鳍缺失导致鲫(Carassiusauratus)和宽鳍鱲(Zaccoplatypus)快速启动能力都显著下降,最大线速度和旋转半径分别下降39% 和62%[5]。本研究发现,尽管背鳍缺失对胭脂鱼幼鱼非稳定游泳能力指标(Ucat、Vmax、Amax、旋转角度和旋转率)没有显著影响(表 1),却导致其捕食者存在条件下的存活率显著下降 35%(图 2)。由此表明,背鳍可能对胭脂鱼幼鱼的反捕食能力具有重要的作用,但其作用并非通过影响其非稳定游泳能力的方式表现出来,具体机制值得进一步研究。先前的研究表明捕食者的口裂大小与猎物的大小密切相关,口裂大小是影响鱼类捕食猎物的一个重要因素[21-23]。因此,胭脂鱼幼鱼高高的背鳍可能是其形态上针对捕食者口裂限制的反捕食策略之一。
声明:本站属公益性没有商业目的的网站,上列文章仅供个人学习参考。本站所发布文章为原创的均标注作者或来源,未经授权不得转载,许可转载的请注明出处。本站所载文章除原创外均来源于网络,如有未注明出处或标注错误或侵犯了您的合法权益,请及时联系我们!
欢迎关注本站(可搜索)"养鱼E线"微信公众帐号和微信视频号"养鱼一线"以及头条号"水花鱼@渔人刘文俊"!
